http://www.photonette.net/
Bird
From Wikipedia, the free encyclopedia
Evolution
There is significant
evidence that birds evolved from theropod dinosaurs, specifically, that birds
are members of Maniraptora, a group of theropods which includes dromaeosaurs
and oviraptorids, among others.[1] As more non-avian
theropods that are closely related to birds are discovered, the formerly clear
distinction between non-birds and birds becomes less so. Recent discoveries in
northeast China (Liaoning Province), demonstrating that many small theropod
dinosaurs had feathers, contribute to this ambiguity.
The basal bird Archaeopteryx,
from the Jurassic, is well-known as one of the first "missing links"
to be found in support of evolution in the late 19th century, though it is not
considered a direct ancestor of modern birds. Confuciusornis is another
early bird; it lived in the Early Cretaceous. Both may be predated by Protoavis
texensis, though the fragmentary nature of this fossil leaves it open to
considerable doubt if this was a bird ancestor. Other Mesozoic birds include
the Enantiornithes, Yanornis, Ichthyornis, Gansus and the Hesperornithiformes,
a group of flightless divers resembling grebes and loons.
The recently (2002)
discovered dromaeosaur Cryptovolans was capable of powered flight,
possessed a sternal keel and had ribs with uncinate processes. In fact, Cryptovolans
makes a better "bird" than Archaeopteryx which is missing some
of these modern bird features. Because of this, some paleontologists have
suggested that dromaeosaurs are actually basal birds whose larger members are
secondarily flightless, i.e. that dromaeosaurs evolved from birds and not the
other way around. Evidence for this theory is currently inconclusive, but digs
continue to unearth fossils (especially in China) of the strange feathered
dromaeosaurs. At any rate, it is fairly certain that avian flight existed in
the mid-Jurassic and was "tried out" in several lineages and variants
by the mid-Cretaceous.

Snowy Owl, Bubo
scandiacus
Although ornithischian
(bird-hipped) dinosaurs share the same hip structure as birds, birds actually
originated from the saurischian (lizard-hipped) dinosaurs (if the dinosaurian
origin theory is correct), and thus arrived at their hip structure condition independently.
In fact, the bird-like hip structure also developed a third time among a
peculiar group of theropods, the Therizinosauridae.
An alternate theory to the
dinosaurian origin of birds, espoused by a few scientists (most notably Lary
Martin and Alan Feduccia), states that birds (including maniraptoran
"dinosaurs") evolved from early archosaurs like Longisquama, a
theory which is contested by most other scientists in paleontology, and by
experts in feather development and evolution such as R.O. Prum. See the Longisquama
article for more on this alternative.
Modern birds are classified
in Neornithes, which are now known to have evolved into some basic lineages by
the end of the Cretaceous (see Vegavis). The Neornithes are split into
the Paleognathae and Neognathae. The paleognaths include the tinamous (found
only in Central and South America) and the ratites. The ratites are large
flightless birds, and include ostriches, cassowaries, kiwis and emus (though
some scientists suspect that the ratites represent an artificial grouping of
birds which have independently lost the ability to fly in a number of unrelated
lineages). The basal divergence from the remaining Neognathes was that of the
Galloanseri, the superorder containing the Anseriformes (ducks, geese and swans),
and the Galliformes (the pheasants, grouse, and their allies). See the chart
for more information.
The classification of birds
is a contentious issue. Sibley & Ahlquist's Phylogeny and Classification
of Birds (1990) is a landmark work on the classification of birds (although
frequently debated and constantly revised). A preponderance of evidence seems
to suggest that the modern bird orders constitute accurate taxa. However,
scientists are not in agreement as to the relationships between the orders;
evidence from modern bird anatomy, fossils and DNA have all been brought to
bear on the problem but no strong consensus has emerged. More recently, new
fossil and molecular evidence is providing an increasingly clear picture of the
evolution of modern bird orders. See also: Sibley-Ahlquist taxonomy.
Bird
anatomy
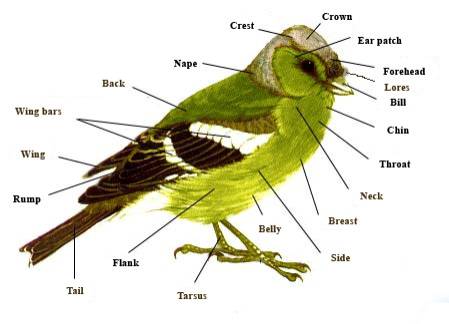
External
anatomy of a typical bird
Birds have a body plan that
shows so many unusual adaptations (mostly aiding flight) that birds have earned
their own unique class in the vertebrate phylum.
The skeleton consists of
bones which are very light. They have large pneumatic cavities which connect
with the respiratory system. The skull bones are fused and do not show cranial
sutures. The orbits are large and separated by a bony septum. The spine has
cervical, thoracic, lumbar and caudal regions with the number of cervical
(neck) vertebrae highly variable and especially flexible, but movement is
reduced in the anterior thoracic vertebrae and absent in the later vertebrae. The
last few are fused with the pelvis to form the synsacrum. The ribs are flattened
and the sternum is keeled for the attachment of flight muscles, except in the
flightless bird orders. The forelimbs are modified into the wings.
Unlike mammals, birds do
not urinate. Their kidneys extract nitrogenous wastes from the bloodstream, but
instead of excreting it as urea dissolved in urine as we do, they excrete it in
the form of uric acid. They also excrete creatine rather than creatinine as in
mammals. Uric acid has a very low solubility in water, so it emerges as a white
paste. This material, as well as the output of the intestines, emerges from the
bird's cloaca. The cloaca is a multi-purpose hole for birds: their wastes come
out of it, they have sex by putting their cloacas together, and females lay
eggs out of it.
Birds have one of the most
complex lung systems of all organisms. Air enters the bird and immediately 75%
of the air bypasses the lungs and flows directly into a posterior air sac which
extends from the lungs and connects with air spaces in the bones and fills them
with air. When the bird exhales the air from the posterior air sac is forced
into the lungs; thus birds receive a supply of air during both inhalation and
exhalation. The gas exchange then takes place in the capillaries. The heart has
four chambers and the right aortic arch gives rise to systemic aorta (unlike in
the mammals where the left arch is involved). Postcava receives blood from the
limbs via the renal portal system. Birds, unlike mammals, have nucleated
erythrocytes, i.e. red blood cells which retain a nucleus.
Sound production is
achieved using the syrinx, a muscular chamber with several tympanic membranes,
situated at the lower end of the trachea where it bifurcates.
The digestive system of the
bird is unique and has a crop for storage and a gizzard for grinding food. Most
are highly adapted for rapid digestion, an adaptation to flight. Migratory
birds have the additional ability to reduce parts of the intestines prior to
migration.
The nervous system,
relative to the bird's size, is actually quite large. The most developed part
of the brain is the one that controls the flight related function while the cerebellum
coordinates movement and the cerebrum controls behavior patterns, navigation,
mating and nest building. A bird’s eyes are developed for taking off, spotting
landmarks, hunting and feeding. Birds with eyes on the side of their head have
a wide visual field while birds with eyes on the front of their heads like owls
have binocular vision and can measure depth.
Most birds have a poor
sense of smell except for the kiwis and vultures. The visual system is highly
developed. Water birds have special flexible lenses, allowing accommodation for
vision in air and water. Some species also have dual fovea. The bird retina has
a fan shaped blood supply system called the pecten. The avian ear lacks
external pinnae but is covered by feathers. The inner ear has a cochlea but it
is not spiral as in mammals.
Wikipedia
http://en.wikipedia.org/w/index.php?title=Bird&action=history
Evolution
There is significant
evidence that birds evolved from theropod dinosaurs, specifically, that birds
are members of Maniraptora, a group of theropods which includes dromaeosaurs
and oviraptorids, among others.[1] As more non-avian
theropods that are closely related to birds are discovered, the formerly clear
distinction between non-birds and birds becomes less so. Recent discoveries in
northeast China (Liaoning Province), demonstrating that many small theropod
dinosaurs had feathers, contribute to this ambiguity.
The basal bird Archaeopteryx,
from the Jurassic, is well-known as one of the first "missing links"
to be found in support of evolution in the late 19th century, though it is not
considered a direct ancestor of modern birds. Confuciusornis is another
early bird; it lived in the Early Cretaceous. Both may be predated by Protoavis
texensis, though the fragmentary nature of this fossil leaves it open to
considerable doubt if this was a bird ancestor. Other Mesozoic birds include
the Enantiornithes, Yanornis, Ichthyornis, Gansus and the Hesperornithiformes,
a group of flightless divers resembling grebes and loons.
The recently (2002)
discovered dromaeosaur Cryptovolans was capable of powered flight,
possessed a sternal keel and had ribs with uncinate processes. In fact, Cryptovolans
makes a better "bird" than Archaeopteryx which is missing some
of these modern bird features. Because of this, some paleontologists have
suggested that dromaeosaurs are actually basal birds whose larger members are
secondarily flightless, i.e. that dromaeosaurs evolved from birds and not the
other way around. Evidence for this theory is currently inconclusive, but digs
continue to unearth fossils (especially in China) of the strange feathered
dromaeosaurs. At any rate, it is fairly certain that avian flight existed in
the mid-Jurassic and was "tried out" in several lineages and variants
by the mid-Cretaceous.
Snowy Owl, Bubo
scandiacus
Although ornithischian
(bird-hipped) dinosaurs share the same hip structure as birds, birds actually
originated from the saurischian (lizard-hipped) dinosaurs (if the dinosaurian
origin theory is correct), and thus arrived at their hip structure condition independently.
In fact, the bird-like hip structure also developed a third time among a
peculiar group of theropods, the Therizinosauridae.
An alternate theory to the
dinosaurian origin of birds, espoused by a few scientists (most notably Lary
Martin and Alan Feduccia), states that birds (including maniraptoran
"dinosaurs") evolved from early archosaurs like Longisquama, a
theory which is contested by most other scientists in paleontology, and by
experts in feather development and evolution such as R.O. Prum. See the Longisquama
article for more on this alternative.
Modern birds are classified
in Neornithes, which are now known to have evolved into some basic lineages by
the end of the Cretaceous (see Vegavis). The Neornithes are split into
the Paleognathae and Neognathae. The paleognaths include the tinamous (found
only in Central and South America) and the ratites. The ratites are large
flightless birds, and include ostriches, cassowaries, kiwis and emus (though
some scientists suspect that the ratites represent an artificial grouping of
birds which have independently lost the ability to fly in a number of unrelated
lineages). The basal divergence from the remaining Neognathes was that of the
Galloanseri, the superorder containing the Anseriformes (ducks, geese and swans),
and the Galliformes (the pheasants, grouse, and their allies). See the chart
for more information.
The classification of birds
is a contentious issue. Sibley & Ahlquist's Phylogeny and Classification
of Birds (1990) is a landmark work on the classification of birds (although
frequently debated and constantly revised). A preponderance of evidence seems
to suggest that the modern bird orders constitute accurate taxa. However,
scientists are not in agreement as to the relationships between the orders;
evidence from modern bird anatomy, fossils and DNA have all been brought to
bear on the problem but no strong consensus has emerged. More recently, new
fossil and molecular evidence is providing an increasingly clear picture of the
evolution of modern bird orders. See also: Sibley-Ahlquist taxonomy.
Bird
anatomy
External
anatomy of a typical bird
Birds have a body plan that
shows so many unusual adaptations (mostly aiding flight) that birds have earned
their own unique class in the vertebrate phylum.
The skeleton consists of
bones which are very light. They have large pneumatic cavities which connect
with the respiratory system. The skull bones are fused and do not show cranial
sutures. The orbits are large and separated by a bony septum. The spine has
cervical, thoracic, lumbar and caudal regions with the number of cervical
(neck) vertebrae highly variable and especially flexible, but movement is
reduced in the anterior thoracic vertebrae and absent in the later vertebrae. The
last few are fused with the pelvis to form the synsacrum. The ribs are flattened
and the sternum is keeled for the attachment of flight muscles, except in the
flightless bird orders. The forelimbs are modified into the wings.
Unlike mammals, birds do
not urinate. Their kidneys extract nitrogenous wastes from the bloodstream, but
instead of excreting it as urea dissolved in urine as we do, they excrete it in
the form of uric acid. They also excrete creatine rather than creatinine as in
mammals. Uric acid has a very low solubility in water, so it emerges as a white
paste. This material, as well as the output of the intestines, emerges from the
bird's cloaca. The cloaca is a multi-purpose hole for birds: their wastes come
out of it, they have sex by putting their cloacas together, and females lay
eggs out of it.
Birds have one of the most
complex lung systems of all organisms. Air enters the bird and immediately 75%
of the air bypasses the lungs and flows directly into a posterior air sac which
extends from the lungs and connects with air spaces in the bones and fills them
with air. When the bird exhales the air from the posterior air sac is forced
into the lungs; thus birds receive a supply of air during both inhalation and
exhalation. The gas exchange then takes place in the capillaries. The heart has
four chambers and the right aortic arch gives rise to systemic aorta (unlike in
the mammals where the left arch is involved). Postcava receives blood from the
limbs via the renal portal system. Birds, unlike mammals, have nucleated
erythrocytes, i.e. red blood cells which retain a nucleus.
Sound production is
achieved using the syrinx, a muscular chamber with several tympanic membranes,
situated at the lower end of the trachea where it bifurcates.
The digestive system of the
bird is unique and has a crop for storage and a gizzard for grinding food. Most
are highly adapted for rapid digestion, an adaptation to flight. Migratory
birds have the additional ability to reduce parts of the intestines prior to
migration.
The nervous system,
relative to the bird's size, is actually quite large. The most developed part
of the brain is the one that controls the flight related function while the cerebellum
coordinates movement and the cerebrum controls behavior patterns, navigation,
mating and nest building. A bird’s eyes are developed for taking off, spotting
landmarks, hunting and feeding. Birds with eyes on the side of their head have
a wide visual field while birds with eyes on the front of their heads like owls
have binocular vision and can measure depth.
Most birds have a poor
sense of smell except for the kiwis and vultures. The visual system is highly
developed. Water birds have special flexible lenses, allowing accommodation for
vision in air and water. Some species also have dual fovea. The bird retina has
a fan shaped blood supply system called the pecten. The avian ear lacks
external pinnae but is covered by feathers. The inner ear has a cochlea but it
is not spiral as in mammals.
Wikipedia
http://en.wikipedia.org/w/index.php?title=Bird&action=history